.jpg "Figure 5 : délétion terminale")
- Pré-requis et Objectifs
-
Cours
- Contenu
- Version Enseignants
-
Version PDF

- Annexes
- Votre Avis
- Ressources Enseignants



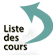
Selon la définition classique, les anomalies de structure sont la conséquence de cassures chromosomiques suivies par un ou plusieurs recollements anormaux. Plus récemment les progrès de la cytogénétique moléculaire ont conduit à proposer trois grands modèles pour expliquer les remaniements du génome humain. La recombinaison homologue non-allélique (non allelic homologous recombination ou NAHR) est le premier mécanisme important de remaniement chromosomique reconnu comme responsable de désordres génomiques. Elle survient pendant la méiose ou la mitose et nécessite deux répétitions segmentaires (low copy repeats ou LCR) ou duplicons, d'une longueur suffisante et avec un haut degré d'homologie, qui agissent comme substrats moléculaires de recombinaison. En raison de leur haut degré d’identité de séquence, les copies non-alléliques de répétitions segmentaires peuvent parfois être alignées en méiose ou en mitose à la place des copies aux positions alléliques habituelles. Ce phénomène, appelé « mésappariement » peut entraîner des remaniements chromosomiques dans les cellules filles. Les copies non-alléliques agissent donc comme des médiateurs (les substrats moléculaires) des recombinaisons homologues. Lorsque les deux répétitions segmentaires se situent sur le même chromosome en orientation directe, une recombinaison homologue non-allélique entre ces derniers entraîne une duplication et/ou une délétion. Si elles se situent sur le même chromosome mais en orientation inverse, une recombinaison homologue non-allélique induit l'inversion du fragment qu'elles encadrent. La plupart des remaniements récurrents, dont l’exemple le plus fréquent est la microdélétion 22q11, résultent de recombinaisons homologues non alléliques: ils partagent une taille commune, des points de cassure groupés, et se retrouvent chez différents individus.
A l’inverse la taille et la localisation des remaniements non-récurrents varie d’un patient à l’autre. Des séquences répétées présentant un haut degré d’homologie (les éléments Alu et LINE par exemple) pourraient induire certaines recombinaisons homologues non alléliques rares responsables de certains remaniements non-récurrents, mais la majorité des remaniements non-récurrents sont expliqués grâce aux modèles de jonction d’extrémités non homologues (non homologous end joining ou NHEJ), qui est un mécanisme de réparation des cassure d’ADN double-brin qui ne nécessite que de très courtes séquences de microhomologie, et celui d’ interruption de la fourche de réplication et commutation de la matrice (Fork Stalling and Template Switching ou FoSTeS). Le mécanisme de FoSTeS est différent de la recombinaison homologue non-allélique et de la jonction d’extrémités non homologues, principalement parce qu’il s’agit d’une voie de remaniement basée sur la réplication qui ne nécessite pas forcément la survenue d’une cassure double brin.
Les anomalies de structure peuvent affecter un chromosome ou deux chromosomes, homologues ou non homologues, parfois davantage ; elles peuvent être équilibrées ou non équilibrées. Les remaniements équilibrées n’entraînent pas de perte ou de gain de matériel chromosomique et n’ont habituellement pas d’effet phénotypique.
Une situation très particulière est celle où la cassure en interrompant un gène au niveau d’un des points de cassure, entraîne une maladie génétique par un phénomène d’haploinsuffisance.
Les remaniements équilibrées peuvent entraîner, lors de la méiose, la formation de gamètes déséquilibrés donnant des zygotes anormaux, ce qui se traduira par la survenue d'avortements ou par la naissance d'enfants porteurs d'anomalies congénitales. Les anomalies non équilibrées peuvent survenir de novo (délétions, translocations non équilibrées, ou autres) ou être la conséquence d’un remaniement parental équilibré.
On différencie les anomalies de structure touchant un seul chromosome des anomalies de structure touchant 2 chromosomes qui impliquent un échange de matériel génomique.
Les délétions résultent d’une cassure chromosomique avec perte du segment distal (délétion terminale), ou de deux cassures sur un même bras chromosomique avec perte du segment intercalaire (délétion intercalaire). Les délétions terminales supposent un mécanisme de restitution d’un télomère pour assurer la stabilisation du chromosome (Figure 5).
Les délétions surviennent le plus souvent de novo. Dans le cas des délétions terminales, une minorité (10 à 15%) résulte de la malségrégation d’une translocation parentale équilibrée ; elles s’accompagnent généralement dans ce cas d’une trisomie pour un autre chromosome (on parle de duplication/déficience).
Les anneaux résultent d’une cassure à chaque extrémité d’un chromosome suivie par un recollement avec perte des segments distaux (Figure 6). Ils sont donc assimilables à une double délétion.

Exceptionnellement un anneau peut dériver de plusieurs chromosomes.
Les chromosomes en anneau sont instables lors de la mitose. Les échanges mitotiques entre chromatides-soeurs engendrent des dérivés complexes avec duplications/déficiences, ce qui complique l’interprétation du phénotype.